Някои аспекти при ухапванията от
пепелянка /Vipera ammodytes/ и усойница /Vipera berus/
Автори: д–р Красимир Христов, Марко Иванов, Георги Русев
За да могат да се дадат по – точни препоръки за първа помощ и последващото лечение, при ухапване от отровни змии в България е добре преди това хората да бъдат запознати с отровния апарат и отровата на двата вида отровни змии, имащи значение у нас – пепелянка /Vipera ammodytes/ и усойница /Vipera berus/
Отровен апарат при пепелянката и усойницата?
![SS.jpg (1807×1204)]()
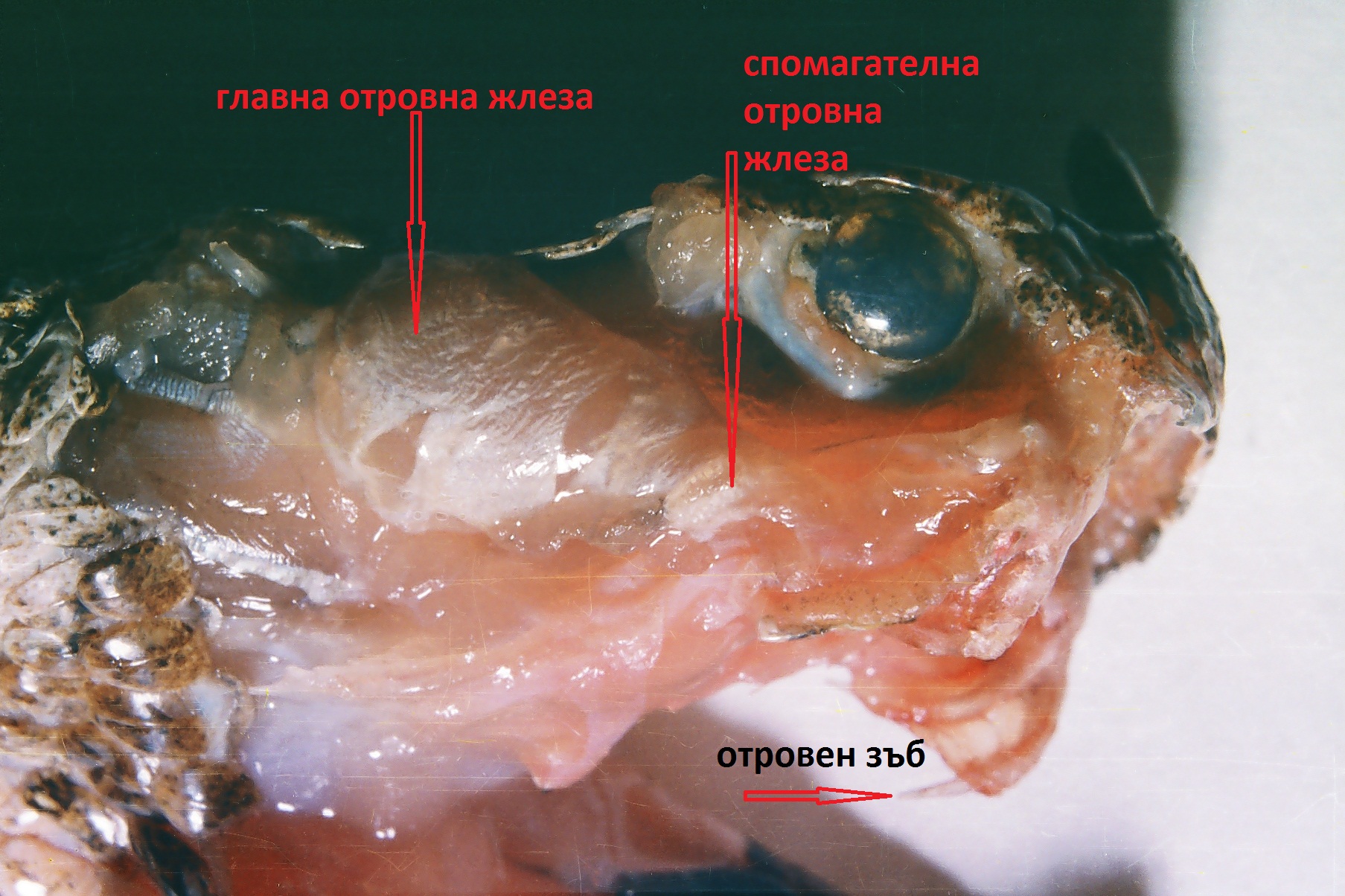
Отровен апарат на пепелянка /V. ammodytes/
Отровния жлеза е двуделна състои се от главна и спомагателна. По морфология е сложна алвеоларна жлеза (представлява видоизменена слюнчена жлеза)
Капацитета на отровната жлеза при пепелянката е много голям.
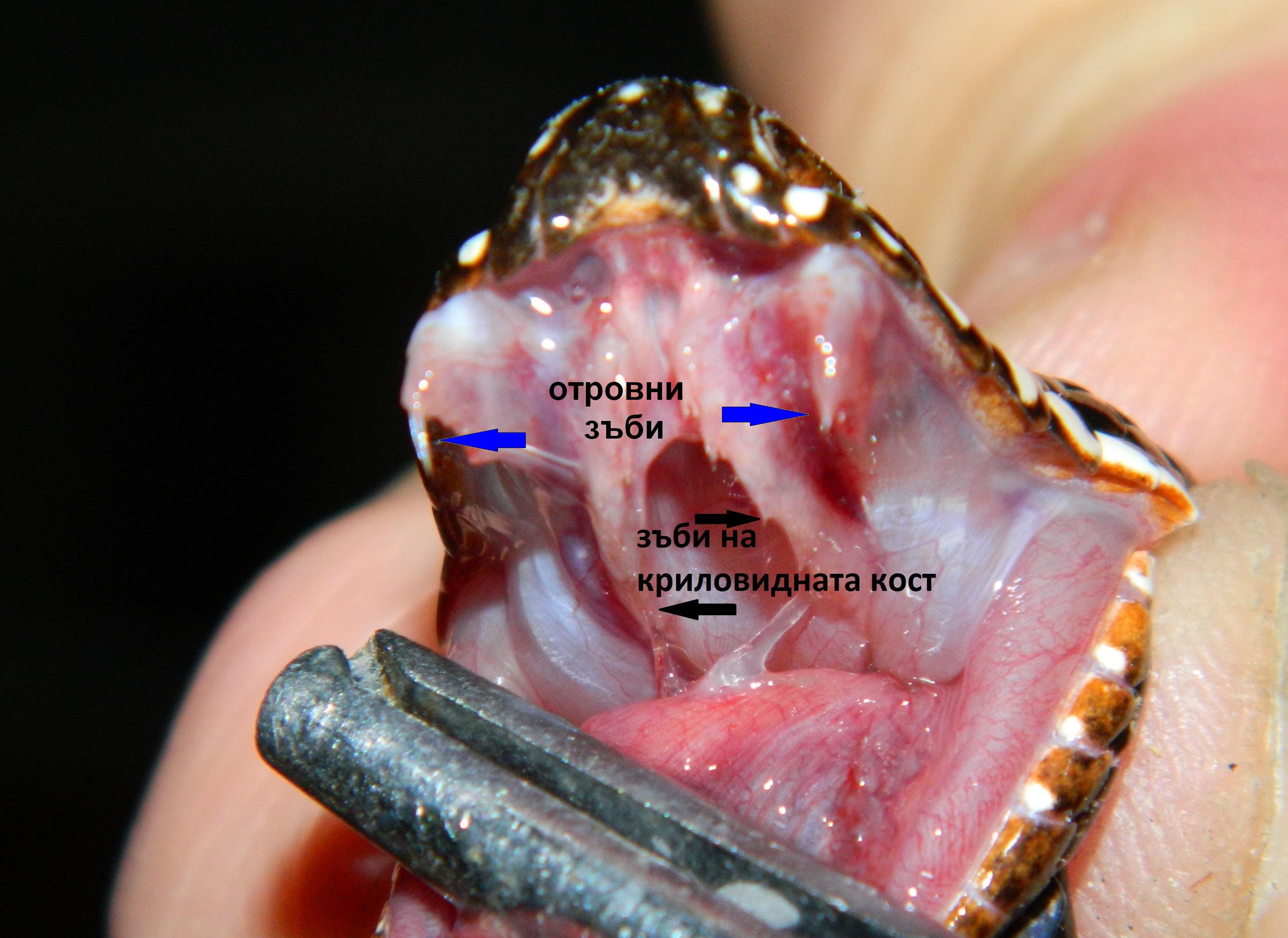
И двата вида са представители на сем. Отровници /Viperidae/т.е. по отношение на зъбите и отровния апарат се категоризират като соленоглифни (Solenogliphia). Това означава, че тези влечуги имат кухи, подвижни отровни зъби, разположени в предната част на горната челюст. Тези зъби са свързани с канали, които водят към отровните жлези. Самата отровна жлеза е двуделна и се състои от главна и спомагателна жлеза. Отделянето на отровния секрет става чрез контракции на дъвкателния мускул (musculusmasseter) и специфичния жлезен мускул (musculus constrictor glandule). Този тип отровен апарат е еволюционно най – напреднал и е характерен за Отровниците /Viperidae/ и Кроталидните змии /Crotalidae/. Отровните зъби са разположени успоредно на горната челюст и при захапване се изправят почти перпендикулярно.
При тези змии освен отровните зъби, по челюстите са разположени и сухи т.е. не свързани с отровни жлези – на горната челюст тези зъби са разположени по криловидните кости и по долната челюст на мандибуларните кости. По горночелюстни кости при солиноглифните змии са разположени само отровните зъби, покрити от лигавични гънки.
![DSCN3674.JPG (2340×1135)]()
Отровните зъби са извити, кухи, крехки и чупливи.
Покрити са с лигавична гънка, която ги предпазва от вредни въздействия.
![DSCN36701.jpg (2304×2472)]()
Когато пепелянката не хапе отровните зъби са разположени успоредно на горната челюст, покрити от лигавичните гънки
Аналогично е положението при усойницата.
![DSCN3645.JPG (4320×3240)]()
Отровните зъби при пепелянка са подвижни и се изправят перпендикулярно на горната челюст при ухапване, но значителна част от дължината им е покрита с лигавична гънка
![DSCN3373a.jpg (3799×2400)]()
Отровния апарат на усойницата /V. berus/ е аналогичен на този при пепелянката
При ухапване отровните зъбите се изправят перпендикулярно на горната челюст, а когато не хапе са разположени успоредно на челюстта. Капацитета на отровните жлези на усойницата е около десет пъти по – малък от този на пепелянката – т.е. при ухапване усойницата е в състояние да отдели приблизително 10 пъти по – малко количество отрова в сравнение с пепелянката. Отровните зъби, обаче, са по – къси.
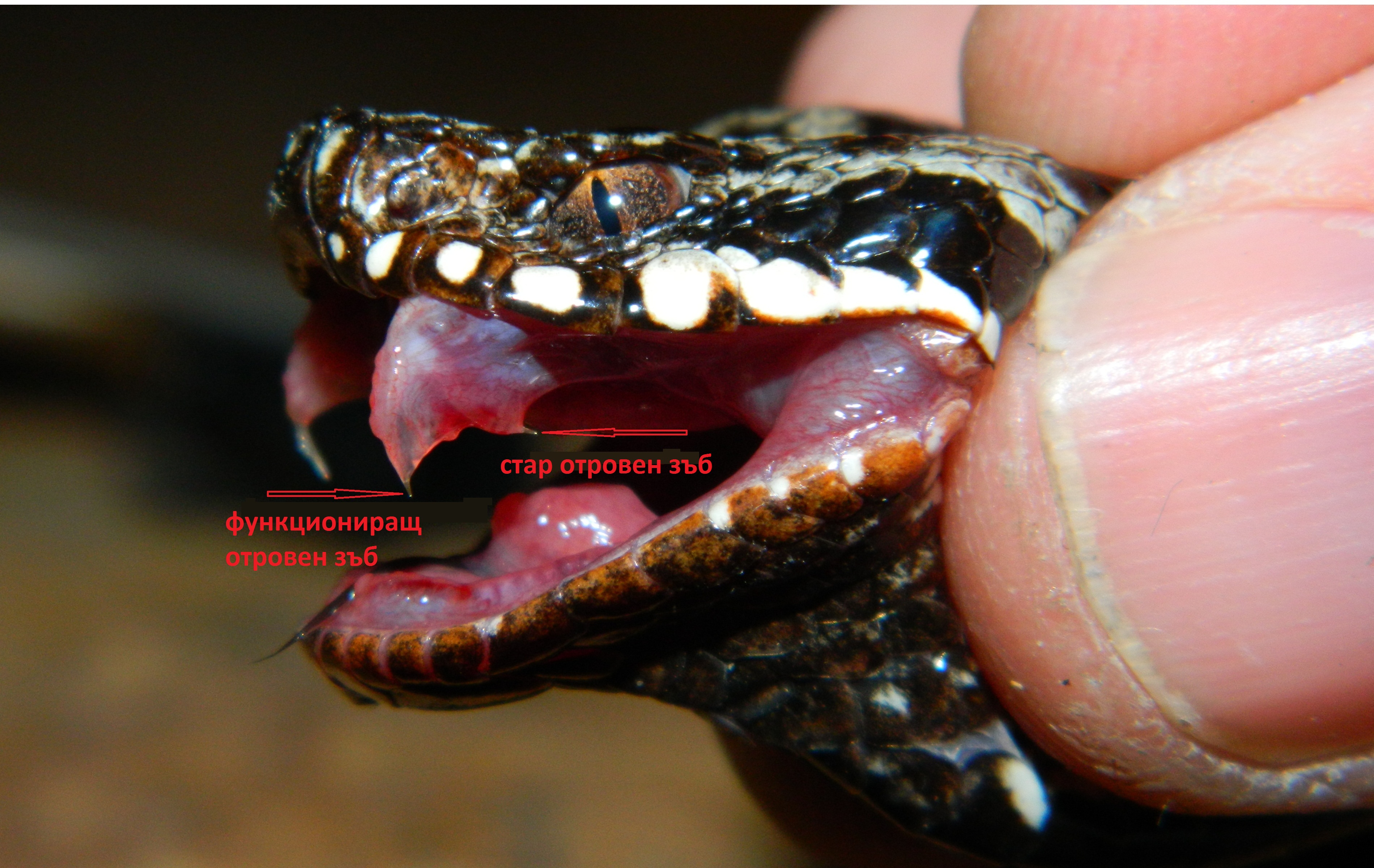
Пепелянките и усойниците често сменят отровните си зъби, които са крехки и чупливи, и понякога остават в ухапаната плячка.
Понякога старите отровни зъби, които не функционират, остават известно време в лигавичната гънка преди да паднат.
Отровата при змиите?
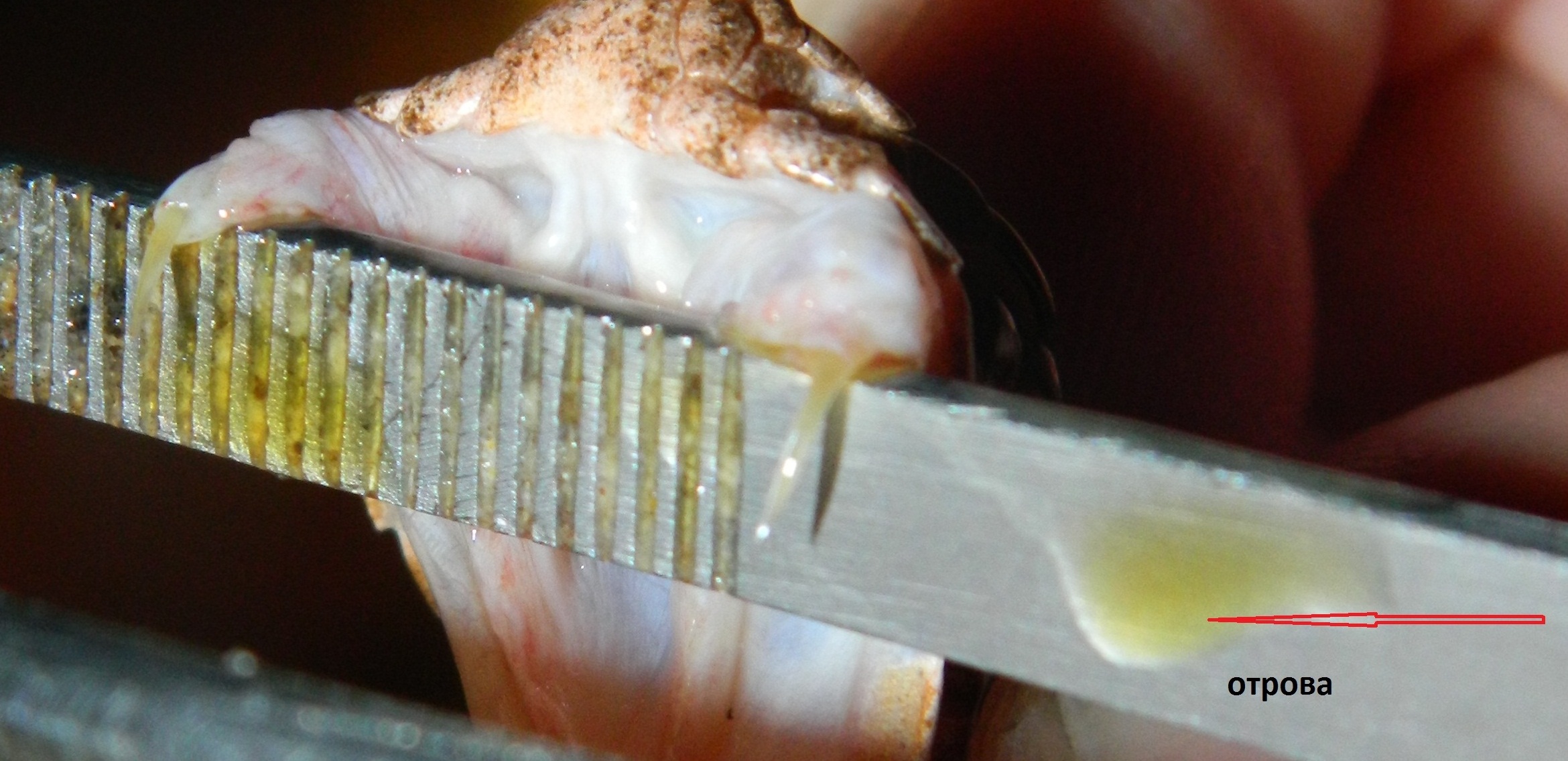
Отровата на тези видове представлява жълта или безцветна, опалисцираща (блестяща) течност, със съдържание на около 30 % сухо вещество около и около 70% вода. Самата отрова представлява смес от пептиди (белтъци) със силно биологично въздействие и високомолекулни съединения, които бъбреците не могат да се отделят в урината. Затова отровата не може да бъде изхвърлена с урината и се открива в тялото на пострадалия до 2 – 3 седмици след ухапването. Повечето токсини обаче губят активността си след 2 – 3 денонощие.
![DSCN3364a1.JPG (2858×2081)]()
Върху горночелюстните кости /os maxilaria/ са разположени само отровните зъби.
На горната челюст сухите не отровни (не отровни) са разположени само върху криловидната кост /ospterygoidea/. Неотровни зъби има и на долната челюст.
![RSCN0782.JPG (1024×768)]()
Отровен зъб на соленоглифна змия.
Изходния отвор на отровния канал се намира на външната страна на зъба, близо до върха.
Променя ли се концентрацията на змийската отрова?
Принципно това не е възможно.
Когато отровата се получава директно, в малки пипети или епруветки, от зъбите тя е с концентрация около 30% сухо вещество и 70 % вода. Понякога при получаването на отровата в петриеви блюда, някои змии отделят по – голямо количество слюнка, поради различни причини, най – вече заболяване, което прави отровата по – водниста и концентрацията на сухото вещество може да падне до 25 – 27 %. Това обаче не се дължи на синтезирането на по – разредена отрова – принципно няма такава. Прецизно получаваните проби винаги са с 30% сухо вещество, от което 90 – 92% са пептиди (белтъчини) и 8 – 10% са различни соли.
Основния фактор, който прави ухапванията от един вид змия (дори и от един и същи екземпляр) много различни едно от друго, не е концентрацията, а количеството инжектирана отрова.
Друг, важен фактор, който влияе върху развитието на самото ухапване е токсичността на самите отрови. Това свойство се определя от т.нар. средна летална доза (LD50), при опитни животни, най – често лабораторни мишки с тегло 18 – 20 g.
Принципно при отделните екземпляри усойницата и пепелянката се наблюдават известни различия в биологичните и имунохимични свойства на отровите, а оттам и в тяхната токсичност. Тези различия могат да бъдат: индивидуални, популационни или подвидови. Най – големи са при двата подвида пепелянки, срещащи се в България. Южната, зеленоопашата пепелянка /Vipera ammodytes montandoni/, отделя отрова с 1,8 – 2 пъти по – висока токсичност, в сравнение с обикновената, червеноопашата пепелянка /Vipera ammodytes ammodytes/ (цвета на опашката, при този подвид е най – често оранжев).
Токсичността на отровата отделяна от българските усойници /Vipera berus/ варира, при пробите взети от отделните екземпляри, но най – често е близка до токсичността на южната зеленоопашата пепелянка (в някои проби е малко по – висока).
Като цяло, потенциалните възможности на отровния апарат на усойницата са около 10 пъти по – ниски в сравнение с тези на пепелянката. Средното количество отрова получавано, чрез мануален масаж, в змиефермите, появили се в страната през 90’ години на миналият век (поддържащи общо взето приемливи условия на отглеждане на влечугите), е около 30 мг суха отрова (отровата получена след изсушаване във вакуум сушилня или ексикатор – нативната, течна отрова не може да се съхранява във вида в който е получена и затова се изсушава веднага след получаването, като количеството суха отрова е основен показател при отровните змии).
![A001.JPG (495×1335)]()
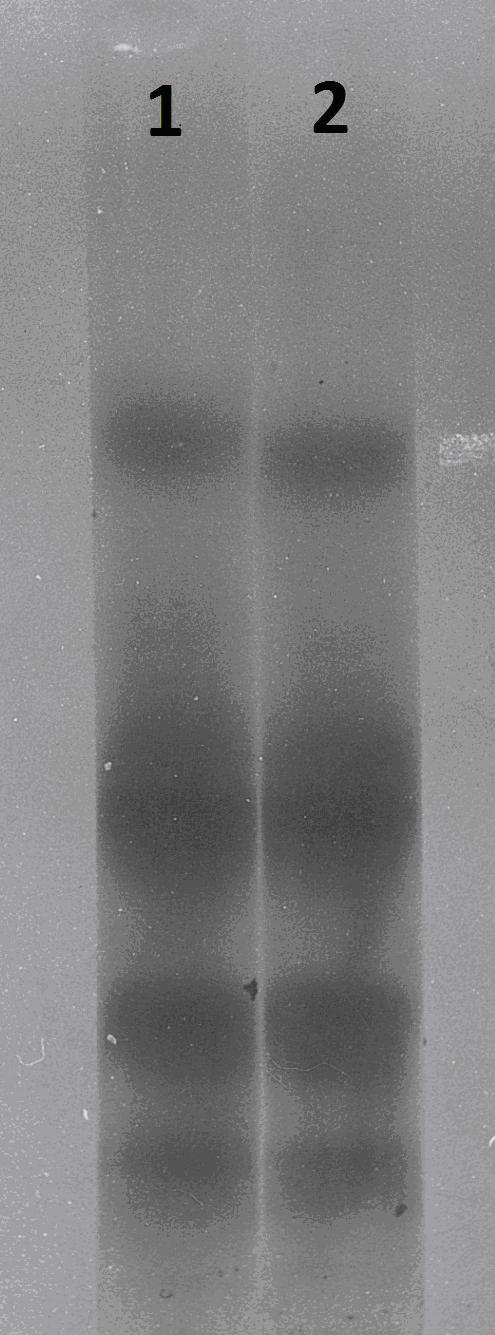
Електрофореза в акрил-амиден гел на отрова от пепелянка
- отрова получена от мъжка южна пепелянка, в пика на нейната активнос (полова активност) 16-часов светлинен ден и температурен градиент 18 - 29ºС;
- отрова получена от същата пепелянка призимуване (6ºС, без светлина).
И двете проби са напълно идентични
Отделни контролни групи пепелянки, в Лабораторията по Херпетология към Тракийския Университет, поддържат средна продуктивност около 52 – 56 мг, като отровата се получава чрез електростимулация.
Максималното количество отрова получено от пепелянка, у нас, е от мъжка пепелянка от Югозападна България, дълга 76 сантиметра, е 130 милиграма (суха маса).
Количеството отрова получавана от усойница е 5 – 7 милиграма (суха маса), (максимално 16 милиграма) – това е около 10 пъти по малко от пепелянката.
Това са показатели за получаване на отрова в лабораторни услови.
Като цяло, при ухапване змиите отделят значително по – малко количество – 1/7 – 1/9 от съдържанието на отрова в жлезите си. Което е в пъти по – малко от полученото, чрез електростимулация или мануален масаж в лаборатория или серпентариум.
Може ли една змия, която е хапала по – често и се е хранила, да отделя по – малко отрова при ухапване?
Като цяло това не е вярно и не почива на здрава логика, защото змиите са в състояние да възстановят почти напълно целия си запас, след получаване на отрова чрез електростимулация, за около месец. Количеството отрова, които се отделят при ухапване са 1/7 – 1/9 от целия запас на змията и се възстановяват много бързо, чрез активен синтез на токсини в отровната жлеза. Някои не токсични съставки идват директно от кръвта и телесните течности на змията (т.е. не са в резултат на активен синтез на жлезата). Противозмийския серум, произвеждан у нас (Бул-Био НЦЗПБ), при кръстосана имунодифузия реагира с кръвния серум на пепелянка, въпреки че кръвния серум не е токсичен.
Количеството отделена отрова зависи най – често от: размера на змията – по – голямата змия има по – големи жлези, респективно повече отрова и състоянието на възбуда в момента на ухапването т.е. до колко застрашена се чувства в дадения момент.
Ако от дадена змия отровата е взета чрез мануален масаж или електростимулация, веднага след това може да се получи ухапване с инжектиране на минимално количество отрова или „сухо” ухапване, но това може да се случи в серпертариум или лаборатория, но не и в природата.
Възможно ли е отровата на дадена змия да е различно токсична през различните сезони?
Принципно при усойниците, в България, не са правени проучвания, но данните получени в Лабораторията по Херпетология на Тракийския Университет (90’ години на миналия век), при пепелянка, показват, че изменението на абиотичните фактори, моделиращи сезонните изменения на климата, по никакъв начин не оказват влияние върху имунологичните, биохимичните и др. фактори на отровата т.е. на нейната токсичност. Накратко казано: пробата отрова взета от зимуваща при 5 – 7ºС, на тъмно пепелянка е абсолютно идентична с пробата отрова от същата пепелянка, в пика на нейната активност (16 часов светлинен ден и температурен градиент 25 – 32СºС). При възрастни пепелянки, поставени при различни микроклиматични условия, не се наблюдават никакви различия в химичния състав и токсичността на отровата. Няма никакви предпоставки за сезонни изменения в отровата при един и същи индивид.
От наши колеги от Института Пастьор, с които сме работили по съвместни изследвания, знаем че аналогично е положението с най – близкия вид до пепелянката вид в Западна Европа – каменарката /Vipera aspis/. Каменарката е западноевропейска отровница, която не се среща в България (границата между ареалите на пепелянката и каменарката се намира по поречието на р. Изонцо, в Италия).
Подобна липса на каквито и да е изменения в биохимичните и токсичните параметри на отровата се съобщава и при други видове отровници и кроталидни змии, така че нямаме основание да очакваме, че може да има някакви сезонни изменения в токсичността на отровата възрастните змии.
Имат ли отрова и могат ли да нанесат ефективно ухапване зимуващите пепелянки и усойници?
Когато при лабораторни условия, преди залягане на зимуване (5 – 7ºС), от дадена пепелянка се вземе отрова, чрез елекртостимулация – практически се получава едно почти цялостно изпразване на отровните жлези – в края на периода на зимуване на същата змия практически не може да се вземе отрова. От това следва, че по време на периода на същинско зимуване (5 – 7 ºС) змиите не продуцират отрова.
Но когато се вземе проба по средата на това зимуване, тази проба е абсолютно идентична с пробата взета в пика на нейната активност. Това показва, че зимуващите пепелянки не продуцират отрова, но ако в жлезите им има такава (това е нормалното състояние), те я запазват в абсолютно не променен вид.
Гореописаното важи за зимуване в строго контролирани условия, но в природата, обаче, не става така. Там зимуващите змии не са във вцепенение, както е описвано по – рано в популярната литература, а те през по топлите зимни дни те излизат от зимовищата и се греят, особено ако няма снежна покривка. През зимата има по – топли дни в които температурата на въздуха достига до около 20ºС, така че е съвсем нормално змиите да синтезират известно количество отрова, ако има някакъв недостиг в жлезите.
Провежданите лабораторни опити показват, че зимуващите пепелянки хапят и безпроблемно убиват лабораторни гризачи, а единият от авторите на тази статия е ухапан ефективно от черна усойница (48 сантиметра.), над циркуса Рибни езера (Рила), при температура на околната среда 6ºС. Това показва, че змиите нямат проблем да нанесат ефективно ухапване при относително по – ниски температури.
Има предположения, че те тогава са в състояние да отделят по – малки количества отрова, но на това не бива много да се разчита!
КАКВО ПРЕДСТАВЛЯВА ЗМИЙСКОТО УХАПВАНЕ
Сухи и ефективни ухапвания
Сухо ухапване– се нарича ухапването нанесено от отровна змия, най - често с отровните зъби, при което не се инжектира отрова.
Сухите ухапвания са напълно реален факт, като при някои видове змии надхвърлят 50%. Практически при сухите ухапвания се наблюдават същите ранички от зъбите, както при ефективно ухапване, но липсват каквито и да е местни или общи симптоми на отравяне. Сухи ухапвания много често се наблюдават в терариумни условия, защото такива влечуги са привикнали с човека и не го възприемат като опасност, но се случват и не рядко в природата. За този факт липсва изчерпателно обяснение, но вероятно е свързан с това, че продуцирането на змийската отрова е свързан с изразходването на големи ресурси на организма и поради това тези животни се стараят да не ги хабят напразно.
При пепелянките има по – голям процент сухи ухапвания, в сравнение с усойниците.това в известна степен може да е обясни с различията в поведението им. При пепелянките се наблюдават по – голям брой спокойни и не агресивни екземпляри, които не хапят ако не са настъпени или сериозно провокирани. При терариумно отглеждани пепелянки много често се наблюдават индивиди, които могат да се вземат е ръка без да са фиксиране (със съответните захвати – с два или три пръста зад врата). При усойниците /Vipera berus/, обитаващи България, това не може да се случи. Дори екземплярите получени като 3 – 4 поколение в терариум са хапливи и нападателни. Поради това сухите ухапвания от този вид се наблюдават значително по – рядко.
Когато говорим за голям брой сухи ухапвания, при терариумно отглежданите змии трябва категорично да изключим змиите отгледани за получаване на змийска отрова. Тези животни преминават не особено приятна за тях процедура – веднъж месечно или на всеки две седмици – свързана с получаването на отровата, а от друга страна се получава хипертрофия на жлезата (особено при елекртостимулация, където липсва травматизъм) и разширяване на канала, което предпоставка за нанасяне на особено тежки ефективни ухапвания.
Сухите ухапвания не се нуждаят от специализирано лечение, достатъчно е раничката да се дезинфекцира за да се избегнат странични инфекции. По принцип тетаносовия бацил /Clostridium tetani/ не обитава устната кухина на змиите, но може да попадне в раничките от външната среда и затова е необходима дезинфекцията със съответните антисептици – общо взето подхода при сухо ухапване от отровна змия е като ухапването от неотровна змия.
Сухите ухапвания всъщност са в основата на множество небивалици, свързани с ухапванията от отровните змии, където се препоръчват най – различни средства като; алкохол, билкови отвари и други небивалици, които са тотално вредни при лечение на ефективно отровно ухапване.
Ефективно ухапване– е такова ухапване, нанесено с отровните зъби на змията, при което в тялото на пострадалия се инжектира някакво количество отрова. Ухапванията се подразделят на: леко, средни и тежко, в зависимост от количеството на инжектираната отрова, като по – правилният подход е да се говори за леко, средно и тежко отравяне, защото основните поражения се нанасят от отровата въведена в организма, а не от механичните травми причинени от зъбите.
Как да различим сухо от ефективно ухапване?
При пепелянката и усойницата това практически е безпроблемно. При ефективното ухапване и при двата вида почти веднага се усеща остра пареща болка в мястото на ухапването.
При усойницата болката е относително поносима, при пепелянката често е по – силна и по – трудно поносима. Малко след това в областта на ухапването се получава бързо разрастващ се оток. По неговото развитие, както и по други общи симптоми може да се съди за каква степен на отравяне се говори – леко, средно или тежко и въз основа на това да се предприеме съответното лечение.
От друга страна дали отравянето ще е по – леко или по – тежко, зависи и от адекватните мерки на първа помощ или липсата на такива.
Най – често целия спектър от клинични симптоми съпътстващи отравянето след змийското ухапване се проявяват през първите 2 часа, има изключения когато първите симптоми на общо отравяне се появяват до 4 – 6 часа, а понякога и след 12 – 18 часа. Известни са случаи когато след известно подобрение е настъпвало ново влошаване след 2 – 3 денонощия. Във всеки случай е необходимо лекарско наблюдение през първите 24 часа и контрол на основните физиологични функции на пациента.
- леко– обикновено се дефинира когато отока не се разпространява в рамките на 30 – 60 мин. – например за посочения период обхваща само пръста и част от китката, няма затруднения в дишането, болезненост при допир около устата или други участъци на кожата (по – често се наблюдава при ухапвания от усойници и понякога при ухапвания от пепелянки);
- средно– ако в рамките на 30 – 60 мин. отока се разраства (за посочения период се разраства над лакътя), наблюдават се симптоми на общо отравяне, болезненост по цялото тяло, понижено кръвно налягане, гадене, повръщане (по – често се наблюдава при ухапвания от пепелянки и рядко при ухапвания от усойници);
- тежко– наблюдава се рядко, при ухапване от едри пепелянки; обикновено симптомите са като при средното, но се проявяват по – силно и по – бързо, като изпреварват развитието на отока – например отока не е обхванал китката, но на 15 – 20 мин. пострадалия не може да стои прав, бързо и бурно развитие на симптомите на общо отравяне, затруднения в дишането, гадене и повръщане (симптоми за средното и тежкото отравяне), по – рядко птозиз, затруднения в говора и други
Усойница /Vipera berus/
![DSCN3869.JPG (4320×3240)]()
Главата на усойницата е относителнопо – малка и по – тясна,в сравнение с главите на другите видове от сем. Отровници(където големите отровни жлези придаваттриъгълно-сърцевидна форма на главата)
![r001-002.jpg (1500×1051)]()
При възрастните меланистичните форми (черни усойници) не се различава отделна шарка на окраската, както и полов диморфизъм.
Новородените меланисти са с нормална окраска (малко по – контрастни) и постепенно с възрастта потъмняват.
Усойницата е по – примитивна и дребна отровница (дължина до около 80 см). В България се среща в планинските райони, най – често на над 800 мнв. Отровата отделяна от тази змия е със средна токсичност, близка до тази на зеленоопашатата пепелянка. Усойницата е общо взето хаплива змия и обикновено хапе, ако бъде приближена на близко разстояние, за разлика от пепелянката. Сравнително рядко нанася сухи ухапвания, но поради изключително ниските възможности на отровния апарат (отделя сравнително малко количество отрова), повечето ухапвания са леки и симптомите могат да отшумят за няколко часа. Понякога макар и рядко има и ухапвания със средна тежест, където е необходимо провеждането на специално лечение и антитоксична терапия. За разлика от пепелянката, където не малък брой ухапвания са по краката, при усойницата са предимно по горните крайници. Това се обяснява с факта, че в планинските части хората ходят с дебели обувки и дълги панталони, които късите отровни зъби на усойницата често не могат да пробият. Ухапванията се случват най – често при бране на горски плодове и катерене по скали и други, като се засягат незащитените части на ръката: длани и пръсти.
Пепелянка /Vipera ammodytes/
![DSCN3814.JPG (4320×3240)]()
Характерна отличителна черта на пепелянката е рогчето на върха на муцуната, но понякога, макар и рядко могат да се открият и екземпляри без рогче.
В България е представена от два добре отличими подвида, с различна токсичност на отровата: обикновена пепелянка /Vipera ammodytes ammodytes/, обитаваща Северозападна България и южна пепелянка /Vipera ammodytes montandoni/ – останалата част на страната.
![DSCN5990.JPG (3240×4320)]()
По – големите отровни жлези на пепелянката придават относително по - плоска относително триъгълно-сърцевидна форма на главата
Средно съотнесена към пропорциите на тялото главата на пепелянката е значително по – голяма от тази на усойницата.
В литературата има описани екземпляри с дължина до 95 сантиметра, най – големия, достоверно, измерен от нас индивид е мъжки с дължина 88 сантиметра.
Това е средна по големина отровница.Значително по – едра змия, в сравнение с усойницата, при еднаква еднаква дължина е 1,7 – 2,5 пъти по – тежка. Отровните зъби са 2 – 2,5 пъти по – дълги. Възможностите на отровния апарат (отровни жлези) са около десет пъти по – големи, от тези на усойницата. Следва се знае, че всяка едра пепелянка има потенциал на отровния апарат да убие възрастен човек. Въпреки това пепелянките нанася голям брой леки ухапвания, след които пострадалите бързо се възстановяват и сухи ухапвания, които въобще не се нуждаят от лечение.
Тази змия е значително по – спокойна, в сравнение с усойницата. Принципно пепелянките избягват човек и ако са близо до убежище се скриват. Ако са далеч от него, обикновено застават неподвижно, като разчитат на защитната си окраска и се надяват да бъдат подминати.
![r001-010.jpg (1500×1051)]()
![DSCN5412.jpg (4320×3240)]()
При обикновената пепелянка /V. ammodytes ammodytes/имаме изразен полов диморфизъм - както мъжките (вляво) са с по – контрастна окраска, която понякога има изразен червеникав оттенък, женските (вдясно) са с по – светла и семпла окраска.
Обикновената пепелянка се среща в Северозападна България и отличава с насочено напред рогче и оранжев или червен връх на опашката. Отделя отрова с относително по – ниска токсичност от южната зеленоопашата пепелянка, но поради големите възможности на отровния апарат, понякога може да нанесе ухапвания съпроводени с тежки отравяния.
![DSCN5340.jpg (4320×3240)]()
![DSCN5467.JPG (4320×3240)]()
При южната пепелянка/V. ammodytes montandoni/също, има изразен полов диморфизъм – мъжките (вляво) са с по – ярка и контрастна окраска, характерната ивица на гърба е по – тъмна; женските (вдясно) са с по – светла и семпла окраска.
Южната пепелянка се среща в равнинните и хълмисти планински части, в цяла България (с изключение на северозападната част на страната). При повечето екземпляри рогчето е насочено нагоре. Цвета на върха на опашката може да варира от ярко зелен до жълтозелен или сивозелен.
Южната пепелянка отделя отрова със сравнително по – висока токсичност, в сравнение с обикновената пепелянка и поради големия капацитет на отровните си жлези (приблизително около 10 пъти по – голям от този на усойницата) е в състояние да нанесе сериозни ухапвания, съпроводени с изключително тежко отравяне. За щастие тези случаи са редки и пепелянките често нанасят леки ухапвания обуславящи слаби отравяния или директно „сухи” ухапвания, при които не се инжектира отрова.
Пепелянките, в България, са значително по – многобройни и разпространени от усойниците и срещите на хората с тях са много по – чести, въпреки това ухапванията са сравнително редки. Това се дължи на факта, че за разлика от усойниците, пепелянките често не хапят, дори ако са приближени на близко разстояние. Ефективни ухапвания най – често се получават ако змията бъде настъпена, притисната или при опит за улавяне.
![DSCN5365.jpg (4320×3240)]()
При някои мъжки зеленоопашати пепелянки характерната ивица на гърба е по – светла, но оградена с по тъмен контур.
Формата на шарката, най – често е ромбовидна, понякога зигзагообразна.
При някои индивиди, ромбовидната шарка преминава в зигзагообразна, в различните участъци на тялото.
Пепелянката е по – едра и с по – дълги отровни зъби, в сравнение с усойницата и може да нанесе ефективно ухапване по долните крайници, през по – тънки обувки или панталони. Поради големите възможности на отровния апарат, някои ухапвания могат да бъдат изключително тежки, където без провеждане на компетентна, масивна специфична антитоксична терапия (лечение с противозмийски серум) може да се стигне до летален изход, особено ако пострадалите са малки деца или хора с хронични заболявания.
![r001-006.jpg (1500×1051)]()
Меланистичните форми при пепелянката се срещат много по – рядко, отколкото черните усойници.
При черните пепелянки има не пълен меланизъм, при който шарката на гърба личи, въпреки, че е по - неясна.
Като цяло ухапванията от пепелянки са много различни едно от друго, в не малко случаи са сухи ухапвания, които не се нуждаят от лечение. Има и леки ухапвания, при които пострадалите могат да се възстановят заедно денонощие, със симптоматично лечение (без серотерапия) сили дори без лечение. Има, разбира се, средни и тежки ухапвания където е необходимо интензивно симптоматично лечение, съчетано с антитоксична терапия (въвеждане на противозмийски серум, които са от решаващо значение за възстановяването на пострадалия.
Повечето пепелянки са спокойни влечуги и не са нападателни или хапливи, но има и агресивни екземпляри, които за щастие се срещат рядко.
Стоящата не подвижно пепелянка, обаче, е трудна за откриване и лесно може да бъде настъпена или притисната. Това е основната причина за ефективните ухапвания в България.
![r001-004a.jpg (1819×1228)]()
![r001-005a.jpg (1819×1228)]()
Вляво: мъжки южни пепелянки – меланистична и нормална окраска
Вдясно: женски южни пепелянки – меланистична и нормална окраска
При черните пепелянки (меланистични форми) се различава характерната окраска, но не и половия диморфизъм, характерен за змиите с нормална окраска
Първа помощ при ефективно ухапване
Главните задачи при доболничната помощ при ухапване от пепелянка или усойница са:
- да се използва всяка възможност за най – бързо доставяне на пострадалия в медицински стационар или болница;
- максимално да се ограничат или ако няма възможност да се отсрочат опасните за живота клинични симптоми на отравянето;
- да се облекчат, с методите на поддържаща терапия първоначалните симптоми на отравяне;
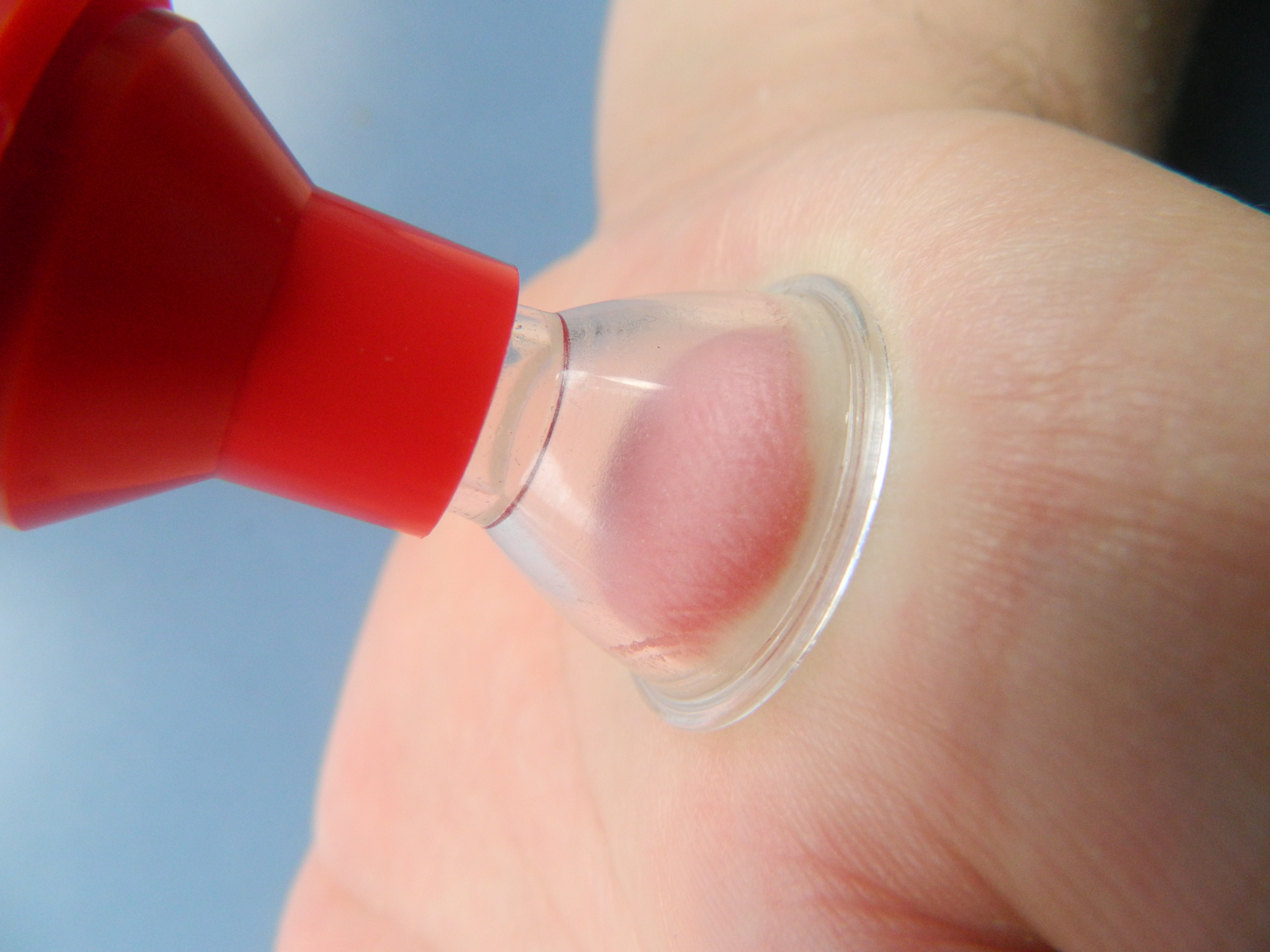
Екстрактор на отрова.
Универсалните екстрактори могат да бъдат използвани не само при ухапване от змии, а и при ужилване от насекоми или убождане от отровни риби.
Съществуват екстрактори с различни конструкции. Добре е да се използват такива модели, които позволяват да се работи с едно движение, дори пострадалия сам да може да извърши изсмукването на отровата.
Да се отчете времето когато е станало ухапването.
Необходимо е да се има предвид, че ухапаните от отровна змия често се намират в състояние на афект. Пред страха от скорошна смърт пострадалите често изпадат в истерия, в някои случаи симулират кома, в някои случаи уплахата може да доведе и до шок. При всички случаи състоянието на афект затруднява отчитането на реалните симптоми и провеждането на лечение. При нужда на пострадалия може да се даде мек дневен транквилант – диазепам или рудотел.
Почистете раната еднократно с влажен тампон, но не я тампонирайте.
Максимално ограничаване (или предотвратяване) на опасните за живота клинични симптоми
За това има само един начин – изсмукване на максимално количество кръв и телесни течности от мястото на ухапването. Тази мярка е единствената която може да намали количеството отрова, попаднала в организма и по този начин да ограничи или при правилни, точни и навременни действия, в някои случаи, буквално да предотврати опасните за живота клинични симптоми.
![DSCN3698.JPG (4320×3240)]()
![DSCN3699.JPG (4320×3240)]()
![DSCN3701.JPG (4320×3240)]()
Обикновено при повечето модели има подробно описание за използването на екстрактора.
При този модел е необходимо само едно движение, след което засмукването става автоматично.
Добре е процедурата по изсмукването да започне до 5 мин след ухапването, като екстрактора се поставя за 30 минути.
Затова, поради тази причина, ще обясним и аргументираме тази мярка подробно.
Всички опити провеждани, от средата до края на ХХ век, опити с по – едри животни (говеда, коне, кози и др.) показват, че извлекът на телесни течност от мястото на ухапването, до 20 – 25 мин. след това, убива лабораторни гризачи (бели мишки). Това се обяснява с факта, че когато не е уцелен магистрален кръвоносен съд (най – често вена) отровата не навлиза веднага в кръвта. Малките кръвоносни съдове, като капилярите, получават спазъм, в досег с нея и се свиват, като отровата най – напред се разтваря в тъканните течности (основно лимфата) и тогава преминава в кръвта. В зависимост от степента на кръвоснабдяване това става за 10 – 20 мин. след момента на ухапване. Точно през това време има възможност, чрез изсмукване най – вече, да се отдели част (или по – голямата част) от змийската отрова попаднала в организма в следствие от ухапването.
Как да стане това?
Описват се случаи на дълбок разрез в мястото на ухапване, някои автори препоръчват дори това да става с нагрято ножче, което е пълна нелепост.
Ефективното ухапването от усойница и особено от пепелянка е силно болезнено, затова някои по – сериозни хирургически експерименти, в такъв момент, биха довели пострадалия до шоково състояние. Още по – голяма глупост са препоръките пострадалия да го извърши собственоръчно.
При ухапване от някои видове змии със сериозни хемокоаголиращи (кръвосъсирващи) свойства на отровата, които не са характерни за нашата фауна, се препоръчва: в мястото на ухапването, между раничките от зъбите, да се направи лек разрез само колкото да се прореже кожата в дистално (отвътре навън) направление с което да се улесни изтичането на кръвта. При ухапванията от пепелянки и усойници общо взето това не е необходимо, защото кръвта сравнително лесно може да се изсмуква от раничките на ухапването. Тази процедура трябва да започне веднага (до около 5 минути) след ухапването и да се провежда интензивно до 30 мин. след ухапването.
Как да се прави изсмукването?
Най – добре е изсмукването в мястото на ухапването да се извършва с т. нар. екстрактор за отрова. Тези устройства които представляват своеобразен вид помпи могат да се поръчват и закупят от много места, не струват скъпо и могат да предотвратят сериозни неприятности и ако имате нужда се снабдете с качествен екстрактор.
При липса на екстрактор за отрова изсмукването може да се извършва и с уста.
Има много не особено компетентни описания, че това е опасно и отровата може да попадне в кръвоносната система чрез раничка в устата. Това в общи линии е напълно нелепо и авторите на тази статия много пъти (над 5 случая) са приемали директно през устата нативна (течна) отрова от пепелянка. Вкуса на отровата на пепелянка варира от сладникаво – тръпчив до леко горчив при различните сборни проби, в зависимост от това дали отровата е жълта или безцветна. Всъщност вкусовите усещания при различните хора са различни затова не претендираме, че изчерпателно сме установили вкусовите качества на тази субстанция, но приетата змийска отрова беше поглъщана, от нас, а не изплювана. При наличието на множество пломби в устата на всеки от нас и язва на дванайсетопръстника при единия, никакви отрицателни въздействия в следствие от приемането на отровата не успяхме да констатираме. Всъщност това не е нещо ново. Много хора преди нас, а вероятно и след нас, са поглъщали змийска отрова без каквито и да е вредни последици за тях. Поради белтъчно – пептидния си характер змийската отрова не е токсична, при поглъщане, за разлика от тази на земноводните, например.
Попадането на нативна (течна) отрова върху някоя повърхностна открита рана върху кожата няма нищо общо с дълбокото змийско ухапване. Това многократно се е случвало с единия от авторите при получаване на отрова на пепелянки и единствения ефект от това е парене, зачервяване или леко подуване на мястото (в много редки случаи). Единствената предпазна мярка която е предприемана в подобни ситуации е измиване на мястото с вода от чешмата. При самото дълбоко ухапване, в подкожните тъкани (при пепелянка може да бъде с дълбочина до около 1,5 см., а при усойницата е по повърхностно, поради по – късите отровни зъби), отровата се инжектира и има възможност за по нататъшната и резорбция в лимфата и оттам в кръвта. При повърхностна рана тези възможности за проникване в организма са редуцирани до минимум
Така че отровата спокойно може да се изсмуква от мястото на ухапване без да съществува някаква опасност за този който го прави. Принципно е добре изсмукваната кръв и телесни течности да се изплюват, но дори да се погълне част от тях не е проблем, много по – опасно е ако отровата остане там където е – в мястото на ухапването. Чисто статистически в токсикологичните справки се отбелязва, че случаите на ухапване, при които е извършена тази процедура, пострадалите пребивават значително по – малко време за лечение в болница, в сравнение с тези които не са я провели. При правилно, навременно приложение на тази мярка, особено при ползване на екстрактор за отрова, може да сведе до минимум (понякога въобще да се прояви) развитието на опасните за живота клинични симптоми.
Отсрочване на опасните за живота клинични симптоми
Ако е спешно е необходимо, на всяка цена, да се отсрочи развитието на опасните за живота клинични симптоми може да се използва стегната превръзка (турникет) над мястото на ухапването. Тази мярка е повече от спорна по отношение на ухапванията на пепелянката и усойницата, защото по – продължителното й използване може да доведе до тежки или понякога, дори, необратими увреждания върху тъканите. Подчертаваме, че турникета има смисъл само като способ за отсрочване на тежките симптоми при ухапване (отравяне) от силно отровни змии и благополучното доставяне на пострадалия (ухапания) до специализирано болнично заведение, където има на разположение противозмийски серум. В противен случай използването на турникета напълно губи смисъл и може да бъде опасно. В световната литература, поставянето на турникет, се препоръчва като отсрочваща мярка при змии отделящи невротоксична отрова – Аспидови/Elapidae/, Морски змии /Hydrophidae/и някои видове Кроталидни змии /Crotalidae/, отделящи невротоксична отрова. Не се препоръчва при ухапвания от Отровници /Viperidae/, към които принадлежат пепелянката и усойницата.
Ние не можем да препоръчаме тази мярка, защото не сме я прилагали. Тези които решат да я използват следва да знаят, че има два основни начина за поставяне на турникет:
- Разхлабен (щадящ)– този турникет спира само отичането на лимфата и е за предпочитане, защото е най – безопасен за ухапания крайник. Поставя се по следния начин: ако имате уред за измерване на кръвно налягане поставете го 10 сантиметра над ухапаното място и напомпте до 55 mmHg. Този турникет може да не се разхлабва в течение на 1 час. При липса на апарат за кръвно налягане, навийте бинт или лента от плат(не гумен маркуч) с ширина около 1,5 – 2 см. Стегнетеe толкова, че под превръзката с натиск да може да се пъхнете един пръст (кутрето). На пристегнатия крайник трябва да се напипва пулс. Този турникет не трябва да се поставя около става.
- Дълбок (артериален)– този турникет се поставя за да се прекрати протичането на кръв и лимфа по ухапания крайник. Не винаги превръзката може да се поставя над ухапаното място, защото, за да се спре артериалното налягане на кръвта, се пристяга около една кост (бедрената кост на крака и лакътната кост на ръката). Такъв турникет може да доведе до сериозни увреждания в тъканите на ухапания крайник и поставянето му е оправдано само при много тежки ухапвания и липса на съответния противозмийски серум, където се налага хирургически да се отстранят част от тъканите в областта на ухапването. Това категорично не е оправдано при ухапване на пепелянки и усойници!
Осигуряване на пострадалия обилно с напитки
С оглед възникването на оток (където се съсредоточава значителна част от течностите на циркулиращата кръв) рехидратирането на пострадалия е от голямо значение.
Съществува схващане, че даването на голямо количество течност на пострадалия (ухапания) се извършва с цел да се стимулира диурезата и да се изхвърлят токсичните съставки чрез урината. Това е погрешно, защото токсините в змийската отрова представляват високомолекулни пептиди (белтъци), които не могат да бъдат отделени през бъбреците. Дори опитите за извеждане на токсините на змийската отрова чрез провеждане на перитониална хемодиализа са били абсолютно безрезултатни. При експерименти с опитни животни змийската отрова се открива до две седмици след въвеждането й.
Рехидрирането на пострадалия е необходимо поради факта, че в място на отока се съсредоточава голямо количество течности от циркулиращата кръв. Поради това е необходимо незабавно рехидратиране на пострадалия за избягване на хиповолемичен шок. Например ако диаметърът на бедрото се увеличи само с 2 см. това може да е причина за съсредоточаване на почти половината течни съставки на циркулиращата кръв в този участък. В следствие от това е необходимо допълнително количество течност, които да компенсират тази загуба, в противен случай кръвта се силно се сгъстява от формените елементи (кръвни клетки).
При осигуряване на пострадалия обилно с питиета (вода, чай – да не са горещи) има един проблем – най – много се нуждаят от течност пострадалите при средни и тежки отравяния, при леките отравяния отока е малък и може да се мине без рехидриране. Само че, обикновено пострадалите при средно и тежко отравяне, с голяма степен на вероятност, ще повърнат всичко което приемат през устата. Повръщането е симптом, в повечето случаи, на тежките отравяния – понякога придобива профузен характер. При тези обстоятелства пострадалия е най – добре да се рехидрира с венозно вливане на разтвори. Тук възниква проблема дали около пострадалия има медицинско или квалифицирано лице, което да постави такава система.
Ранни симптоми
Появилите се ранни симптоми се лекуват със симптоматично средства, като се записва времето им на поява. Болката при ухапване на пепелянка понякога е изключително силна, малко по – поносима е при ухапване на усойница, но и в двата случая е добре да се дават болкоуспокояващи, но да се избягват такива като аспирина, които могат предизвикват повръщане. Най – добре е ползването на аналгин и парацетамол. При развитие на остро отравяне е препоръчително нищо да не се дава през устата, защото лесно може да бъде повърнато.
Гадене и повръщане са ранните симптоми на сериозното отравяне. Пострадалия следва да се постави да легне на една страна, главата да се отпусне леко надолу. Ако повръщането е с профузен (непрекъснат) характер следва да се даде една таблетка хлоразин (хлорпромазин).
Алергични реакции към змийската отрова
Известни са много случаи на силни алергични реакции към змийската отрова, включително при ухапвания на пепелянки и усойници. Тези реакции са по – чести при хора получили змийски ухапвания по – рано или работещи със суха змийска отрова. Незабавната, силна алергична реакция е известна като анафилаксия. Късната алергична реакция е известна като серумна болест.
Признаците на анафилаксията: падане на кръвното налягане; оток на лигавицата на устата, устните и лицето; спазми на бронхите; коремни колики; повръщане; остра слабост и други. Главата на пострадалия трябва да се отпусне надолу, да се даде на пациента антиалерзин, алергозан 2 таблетки или фенкарол. При бързо прогресираща анафилаксия да се инжектира 0,5 % разтвор 1 ml адреналин подкожно (ако е възможно венозно) и инжективни форми (ампули) алергозан, димедрол, антиалерзин, супрастин).
Не бива да се бъркат симптомите на алергия с тези на отравянето, в следствие на змийско ухапване, въпреки някои общо приети погрешни схващания. Те са различни и в голяма степен се лекуват по различен начин.
![bscap0002.jpg (720×576)]()
![bscap0003.jpg (720×576)]()
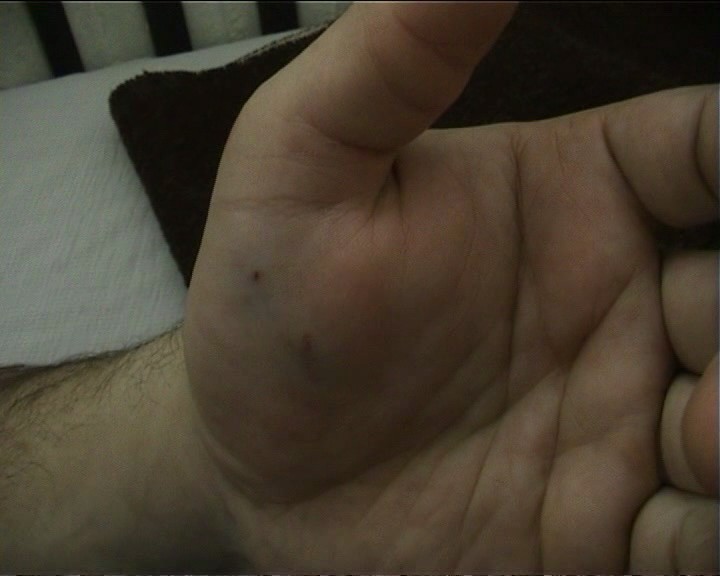
Тежко ухапване от пепелянка, съпроводено със сериозно отравяне на организма..
Ясно се виждат следите от двата отровни зъба, като голямото разстоянието между двете ранички показва, че е нанесено от едра змия.
Отока, все още не е започнал да се развива, а са налице симптоми на общо отравяне (около 20 минути след ухапването): гадене, повръщане, затруднения в дишането, пострадалия не може да стои прав.
Тези характерни белези (само от отровните зъби), обаче, се наблюдават при сравнително малък процент ухапвания. Най – често имаме от една до няколко ранички, като някои са нанесени от отровните зъби, други от неотровните зъби. Не е задължително да са подредени под някаква форма.
Успешно възстановяване след десет дни лечение.
Ухапвания в природата
За нашите географски ширини ухапванията от змии, най – често, се случват по крайниците.
В тази насока следва да се препоръчат следните спешни действия:
- Ухапания крайник може да се обездвижи като при счупване;
- Ако ухапването е по ръката фиксирането трябва да се направи така, че следите от зъбите да са по – ниско от сърцето; да се върви бавно като се спира и почива на всеки 5 минути; ако се използва стегнат турникет е необходимо да се освобождава за 1 минута, на всеки 10 минути; затова трябва да се спира и почива, в легнало положение, 2 минути;
- При ухапвания от пепелянка и усойница кръвта от мястото на ухапването следва да се изсмуква, най – често, без допълнителни разрези, най – добре с екстрактор;
- При нашата практика веднага след ухапването се инжектира 40 – 80 мг метилпреднизолон. Ако преди това пострадалия има предишно ухапване или е работил със суха отрова, следва веднага да се вземе съответния антихистаминов препарат. В България най – често като антихистамини се използват: алергозан, клемастин, антиалерзин и други. Антихистаминовите средства имат страничен седативен (сънотворен) ефект; това е неблагоприятно при респираторна (дихателна) недостатъчност. При хора, получили предишни ухапвания или работещи със суха отрова следва да се прилагат незабавно поради опасност от анафилактичен шок;
- Ако има под ръка противозмийски серум може да се инжектира на няколко пъти, разреден по възможност във физиологичен разтвор, по Безредка;
- Ние лично сме въвеждали серум в мястото на ухапване от пепелянка и ефекта от това е много положителен;
- При спешни случаи когато серум трябва да се въвежда в извън болнична обстановка това трябва да става задължително с инжектиране на 50 – 100 mg метилпреднизолон, след предварително приемане на антихистаминов препарат (алергозан, фенозепам, супрастин)
При всички случаи категорично да не се употребява алкохол!
СЪКРАТЕН РЕД ДЕЙСТВИЯ НА ПЪРВА ПОМОЩ
ПРИ УХАПВАНИЯ ОТ ОТРОВНИ ЗМИИ
(при ефективно ухапване)
- Отчетете времето на ухапването;
- Успокойте пострадалия, при необходимост му дайте мек дневен транквилант – диазепам или рудотел;
- Сложете го да легне и не му позволявайте активни движения;
- Изтрийте мястото на ухапване с влажен тампон, за отделяне на евентуални остатъци от отрова;
- Отделете от раничките максимално количество кръв – най – добре е това да става с екстрактор за отрова, но ако не е наличен такъв това може да се прави и с уста, без никаква опасност за този който го извършва, изсмукваната течност следва да изплюва. Тази манипулация има смисъл да започне до 5 -6 минути след ухапването. Принципно отровата на двата подвида пепелянки и усойницата няма особено влияние върху кръвосъсирването и в почти всички случаи кръвта може да се изсмуква директно от раничките. При нужда могат да бъдат направени два не много дълбоки разреза (прорязващи само кожата), които да улеснят отделянето на кръвта от ухапаното място. Разрезите се правят в надлъжната ос на ухапания крайник, в дистална посока към мястото на ухапване и трябва да са с дължина около 1 сантиметър. Екстракторът на отрова се поставя за около 30 мин или се изсмуква с уста;
- Обездвижете ухапания крайник във функционално положение, като мястото на ухапването да се намира под равнището на сърцето;
- Осигурете в помещението достъп на свеж въздух;
- При възможност измерете пулса, кръвното налягане и температурата на пострадалия;
- Обезпечете пострадалия с обилно питие, но да не е горещо, ако не повръща;
- В случай, че симптомите не прогресират бързо, по правило може да се ограничите само до тези мерки;
Ако в течение на 10 – 30 минути се появят нови симптоми на отравяне и прогресират старите е необходимо:
- да се инжектира на пострадалия 30 – 90 мг метилпреднизолон, 25 – 50 mg супрастин (или алергозан, димедрол, антиалерзин, в аналогични дози). При поява на силни болки дайте аналгин или парацетамол;
- поставете венозна капкова система с глюкоза или физиологичен разтвор (най – добре Хартманов разтвор) (тази операция следва да се извърши от лице с медицинска подготовка) и подгответе съответния противозмийски серум за въвеждане;
- при поява на симптоми на свръхчувствителност и анафилактичен шок (рязко падане на кръвното налягане, оток на лигавицата на устата, нарушено дишане, болки в корема, повръщане) е необходимо да поставите главата на пострадалия ниско отстрани. Инжектирайте 0,5 – 1 ml адреналин, подкожно (ако е възможно венозно). Усилете вентилацията в помещението.
Л Е Ч Е Н И Е
Интоксикацията (отравянето) след ухапване на отровна змия предизвиква 10 – 12 клинични симптома, които могат да се проявяват по няколко заедно, внезапно или постепенно да следват един след друг. Лекуващият лекар трябва да ги диагностира с помощта на клинични и лабораторни методи и да има възможност да извършва поддържаща терапия.
Типични клинични симптоми при отравяне със змийска отрова :
- преразпределение на елементите на циркулиращата кръв;
- хиповолемия (намаляване количеството на циркулиращата кръв) и понижаване на кръвното налягане;
- загуба на плазмени белтъци;
- разрушаване на тромбоцитите (кръвни плочки);
- изменения в количеството и качеството на еритроцитите (червените кръвни клетки), вкл. хемолиза;
- нарушения в стените на кръвоносните съдове;
- некроза на тъканта;
- оток на белите дробове;
- нарушения на сърдечната дейност;
- неврологични нарушения.
Засега няма химически, физически и хирургически средства, които непосредствено да оказват влияние върху измененията, предизвикани от змийската отрова в организма освен противозмийски серум.
Поради това отсъствието на съответния противозмийски серум, при ухапване от всяка опасна отровна змия, поставя лекуващия лекар в сложно положения.
Мястото на ухапването следва да се обработи с антисептичен разтвор и да се покрие със стерилна марля. Да не се привързва. Ухапания крайник следва да се постави във функционално положение.
При ухапване по дланта или пръст на ръката, пръстите трябва да се сгънат под ъгъл близък до 90º в метафалангиалните стави, а в междуфалангиалните леко сгънати. При ухапване по крака, крайника се следва да се постави под ъгъл близък до 90º.мястото на ухапването трябва са бъде под равнището на сърцето през първите 24 часа. Стегнатото бинтоване може да предизвика затруднено изхранване на тъканите (исхемия).
Хиповолемичен шок
Така наречената хиповолемия (хиповолемичен шок) е свързан с намаляване количеството на циркулиращата кръв. Това състояние винаги се съчетава с хипотония (понижени кръвно налягане). Този симптом е свързан с развитието на голям оток след ухапването и е следствие от задържането на в голямо количество течности в самия оток.
Такъв шок се наблюдава 6 – 12 часа след ухапването. Обаче при бързо развиващ се оток хиповолемичен шок може да настъпи за 1 – 2 часа. Например: увеличението на бедрото с 2 сантиметра може да е следствие от задържането на почти от половината от течностите на циркулиращия кръв.
Обикновено това е основната причина за ранната смърт при едри Отровници и Кроталидни змии. При ухапвания от пепелянки и усойници се проявява в по – малка степен, но все пак е един основните симптоми, при средни и тежки отравяния, в следствие от ухапвания на тези два вида.
Развитието на коронарна недостатъчност при хиповолемичен шок води до смърт при отсъствие на бързи терапевтични мерки. При ухапвания от пепелянка и усойница най – често не се налага интубиране на пострадалия и подаване на кислород. Добре е, обаче, да се провери рН на кръвта и при рН под 7,35 пациента се намира в състояние на респираторна ацидоза. Основна мярка е да се постави венозна система – при хиповолемичен шок албумина (5%) е за предпочитане пред 5% глюкоза или 0,9% NaCl. Ако липсват възможности за вливане на албумин, може да се вливат другите разтвори или Хартманов разтвор. Ако налягането в белодробната артерия е по – ниско от 11 torr или в централната вена е по – ниско от 9 torr (12 см Hg), разтворът следва да се влива със скорост 20 ml/min в продължение на 10 мин. бързото повишаване на налягането в белодробната артерия или централната вена може дa способства за развитието на оток на белите дробове, затова е добре да се въвеждат препарати изопреналин изопренол хидрохлорид). Ако въвеждането на течности не дава достатъчно бърз ефект може да се използват малки дози адреналин или изопреналин.
Използването на кортикостероиди (метилпреднизолон, хидрокортизон) при хиповолемичен шок е дискусионен въпрос. Тези препарати нямат вазоконстрикторно действие и не свиват кръвоносните съдове на периферията. Кортикостероидите на снемат действието на змийската отрова върху организма. Единствената им роля при шоково състояние е да увеличават минутния обем на сърцето. Почти всички терапевти работели върху лечението на тежки отравяния, причинени от опасни змии, дават положителна оценка на тези препарати. При ухапвания в лабораторията ние използваме метилпреднизолон като незабавна мярка на първа помощ и по всяка вероятност употребата им при стационарно болнично лечение е оправдана.
Всички гореспоменати мерки при хиповолемичен шок са слабо ефективни или дават краткотраен ефект без въвеждане на противозмийски серум. Серумът трябва да бъде въвеждан незабавно при всички случаи на хиповолемичен шок, тъй като само симптоматичните лечебни мерки не могат да дадат стабилен ефект.
Противозмийският серум обаче не влияе върху физиологичните процеси в организма и въвеждането му трябва да се съчетава със симптоматично лечение.
Други симптоми
Развитие на хемолиза (разрушаване на червени кръвни клетки) – не се наблюдава при повечето ухапвания от усойници и пепелянки, но е полезно отчитането на хематокрита (съотношението между обемите на кръвната плазма и кръвните клетки, нормалното съотношение е 60:40 или 55:45). Полезни са също така и изследвания за протеинурия и хемоглобинурия.
Неврологични нарушениякато: птозис парези по крайниците и болки в междуребрените мускули при вдишване и издишване се наблюдават рядко, въпреки че, в отровата и на усойницата, и на пепелянката се съдържат невротоксични компоненти. Действието на тези невротоксини, се проявява слабо на фона на другите симптоми на отравянето.
Некрозина тъканите в областта на ухапването, при пепелянки и усойници, се наблюдава сравнително рядко, най – често като следствие от неправилни хирургически действия.
Като правило гореописаните симптоми бързо отшумяват при правилна антитоксична терапия (серотерапия) и подходящо симптоматично лечение.
Л Е Ч Е Н И Е С П Р О Т И В О З М И Й С К И С Е Р У М
( С Е Р О Т Е Р А П И Я )
![DSCN5577.JPG (3240×4320)]()
Противозмийски серум на Бул–Био НЦЗПБ има активност срещу отровата на усойницата и пепелянката
Методиката за получаване на противозмийски серум е създадена в края на 19 век (около 1895 г.) от доктор А. Калмет в Пастьоровия институт. В наши дни много институти използват съвременни модификации на разработената от Калмет методика. Противозмийският серум се получава предимно от коне, както е в България в момента, (по – рядко от кози или овце) които се имунизират постепенно с нарастващи дози отрова. Получените антитела остават в кръвния серум и се пречистват чрез хроматографски методи. През последните десетилетия противозмийските серуми не съдържат пречистени антитела, а само части антитела, така наречените Fab – фрагменти (fragment antigen binding-фрагмент който се свързва с антигена). Fab – фрагментите са получават при хидролиза (разделяне) на пречистените антитела с фрагмента пепсин.Така се получават (Fab)2 фрагменти-два Fab фрагмента свързани с дисулфидна връзка. По този начин се постигат по – високи антитоксични концентрации и се намалява вероятността за алергични реакции при употреба на противозмийски серум.
Противозмийските серуми биват моновалентни, предназначени за лечение на ухапване от един вид змия, или поливалентни, предназначени за лечение на ухапвания от няколко вида змии (които понякога са от различни семейства), характерни за даден район.
В България серум се получава чрез хиперимунизация с отрова на пепелянка. Поради изразената си параспецефична активност този серум обезврежда както отровата на пепелянката, така и тази на усойницата.
Антитоксична активност на противозмийския серум
Под антитоксичност на противозмийския серум се разбира неговата способност да обезврежда змийски отрови. Тези субстанции са биопродукти, при които различните партиди имат различна антитоксична активност, като някои са с по – висока активност, други с по – ниска. Затова при тях се определят минимални антитоксични нива, под които антитоксичната активност на серума не трябва да пада.
Според изискванията на Европейската фармакопея противозмийските серуми, в Европейския съюз, трябва да имат следните минимални нива: 1 ml серум трябва да обезврежда не по – малко от:
100 LD50 Vipera ammodytes(пепелянка);
100 LD50 Vipera aspis(каменарка);
50 LD50 Vipera berus (усойница);
50 LD50 Vipera ursinii(остромуцунеста усойница).
При такава активност 5 ml серум (1 ампула) ще обезврежда около 1,5 mg суха отрова от Усойница, около 3 – 4 mg отрова от Южна пепелянка и около 6 – 7 mg от Обикновена пепелянка. Това са приблизителни, но ориентировъчни стойности за активността на една ампула противозмийски серум (5 ml).
Българските противозмийски серуми се стандартизират към 100 АЕ (антитоксични единици) в една доза около 5 ml, по методиката разработена от проф. д-р Р. Манахилов, като активността им е почти идентична с тази на европейските.
Някои партиди серуми имат по – висока антитоксична активност от стандартизираните, но те обикновено се смесват с партиди с по – слаба активност, за да може всички количества пуснати на пазара да покриват необходимите минимални антитоксични стойности.
Ако се изхожда от минималните необходими антитоксични нива, може да се изчислят нужните приблизителни количества за ефективно обезвреждане на отровата при съответно по – леки, средни и тежки ухапвания (отравяне).
Не винаги е лесно да се определи какво количество отрова отделя една змия при ухапване. Това зависи от вида, размерите, до каква степен влечугото се чувства застрашено и други. Повечето автори изследващи този проблем определят, че отровните змии отделят от 1/10 до 1/7 от отровата, която се намира в отровните жлези. Като знаем, че при отделяне на отрова чрез електростимулация, змиите отделят около 80% от запасите си. На тази база може да се определи приблизителният потенциал на отровните жлези, при отделните екземпляри усойници и пепелянки. От потенциала на отровните жлези, може да се прогнозира количеството отрова при ухапване – разбира се, твърде приблизително.
При усойницата, чрез електростимулация, се получават 7 – 12 mg отрова (суха маса). При тези възможности възрастните усойници биха били в състояние да отделят 1 – 3,5 mg при ухапване (суха маса). За неутрализирането на такова количество отрова биха били необходими 1 – 2 ампули (дози) серум, респективно 5 – 10 ml серум, покриващ изискванията за минимална антитоксична активност на Европейската фармакопея.
При пепелянките потенциала на отровните жлези е много по – голям (около 10 пъти). През 90’те години на миналия век, съответно в по – добрите змиеферми, чрез мануален масаж се постигаше около 30 mg средно. В Лабораторията по Херпетология отделни групи опитни животни достигаха среден добив до около 50 mg, чрез електростимулация. При такъв потенциал на отровните жлези може да смятаме, че средно големите пепелянки могат да отделят средно приблизително 8 – 9 mg отрова (суха маса). За инактивирането на такова количество отрова биха били необходими около 10 – 15 ml серум (т.е. около 2,5 – 3 ампули), при южна пепелянка. При обикновена пепелянка, за неутрализирането на същото количество отрова ще са необходими 1,5 – 2 дози (7,5 – 10 ml серум), поради по- ниската токсичност на отровата.
Тези данни се определят въз основа на средна групова продуктивност от пепелянки, отглеждани при лабораторни условия. При ухапване, обаче, биха се проявявали индивидуалните особености на отделните екземпляри, а не средни показатели за цяла опитна група змии. От индивидуалните проби получавани в Лабораторията по Херпетология и в терариума на Тракийско Херпетологично Дружество знаем, че от отделни едри пепелянки (най – често мъжки индивиди), могат да се получат много по – големи количества отрова 70 – 80 mg отрова (суха маса). Такива екземпляри спокойно могат да отделят 12 – 14 mg суха отрова, при ухапване. За инактивирането на подобни количества отрова ще са необходими 5 – 6 дози ( около 25 – 30 ml) за отровата на Южната пепелянка и 2 – 3 дози (около. 10 – 15 ml) за отровата на Обикновената пепелянка.
Максималното количество отрова, получавано в България, е 130 mg и е от мъжки екземпляр, намерен в Югозападна България. Такива едри мъжки пепелянки, които се срещат доста рядко, биха могли спокойно да инжектират при ухапване 21 – 23 mg отрова. За неутрализирането на подобно количество отрова ще са необходими 7 – 8 ампули за Южна пепелянка и 3 – 4 ампули за Обикновена пепелянка.
Тези изчисления са твърде приблизителни, но все пак биха могли да ориентират за необходимите количества серум с минимална, стандартна активност за обезвреждане на змийската отрова попаднала в организма, след леки, средни и тежки ухапвания.
Трябва да се има в предвид, че титрацията на противозмийския серум, с която се измерва неговата активност, по – голяма част от опита се извършва in vitro. Реакцията антиген – антитяло се преминава в термостат при 37ºС в няколко милилитра буфериран физиологичен разтвор. При реални условия където змийската отрова (антигените) и противозмийския серум (антителата) са разредени в няколко литра телесни течности реалната антитоксична активност е относително по – малка.
Тест за поносимост към противозмийски серум
Противозмийския серум представлява пречистени антитела (съвременните серуми се състоят от фрагменти на антитела – Fab – фрагменти) от конски кръвен серум.
Поносимостта към кръвния конски серум се определя най – често чрез интракожна (подкожна) и по – рядко интраконунктивна проба. Най – добре е да се прочетат препоръките в инструкцията на съответния серум.
Интраконунктивните проби често дават положителни резултати. При интракожна (подкожна) проба много пациенти, които преди това са имали множество алергични реакции към различни антигени дават отрицателен резултат. Повечето от тези пациенти получават анафилактичен шок дори при въвеждане на 1% от наличната доза на серума. Късни алергични реакции, като серумна болест, въобще не могат да бъдат открити чрез теста за хиперинтензивност.
Най – чести 1:100 или 1:10 разтвор на серум се въвеждат интракожно докато се образува подутина с диаметър около 2 mm, най – често на ръката, на която не се поставя венозна система. Теста се проверява след 15 мин. Ако няма зачервяване и подутината не се е увеличила с 1 сантиметър, пробата се счита за отрицателна. Ако резултатите са съмнителни теста се проверява след още 15 мин.
Положителния тест не се явява абсолютно противопоказание за използването на противозмийски серум. При ефективно ухапване на опасна отровна змия, където има реална опасност за живота на пострадалия, серума следва да се въвежда независимо от чувствителността към него. Това трябва да се направи след предприемането на съответните противошокови мерки.
Въвеждане на серума
Най – добрия начин за въвеждането на серума е венозно чрез система, разредена с 5% разтвор на глюкоза или Хартманов разтвор за 30 – 60 минути.
Серумът може и да се инжектира венозно със скорост 5 ml/min. При този начин на въвеждане симптомите на свръхчувствителност най – често се развиват до 10 – 15 минути след инжектирането на противоотровата.
Противозмийският серум също така може да се инжектира мускулно под формата на няколко дълбоки инжекции във фронталната (предната) или латералната (страничната) повърхности на бедрото или в големите мускули на ръката. Този начин, по метода на Безредка (поставяне на няколко разредени с физиологичен разтвор части от една доза (ампула), през определен интервал от време) се предпочита от лекарите в България. При сериозно отравяне той не осигурява бърза антитоксична терапия и забавя обезвреждането на проникналата в организма отрова. Дори в страни от Западна, Централна и Северна Европа, които се обитават от по – малко опасни видове отровни змии, сравнени с България, се препоръчва въвеждането на противозмийския серум в разреден вид чрез венозна система. Този метод е приет в целия свят и без съмнение осигурява най – добри възможности за антитоксична терапия.
Понякога 1/3 от серума (разреден с физиологичен разтвор) може да се инжектира в мястото на ухапването. Някои лекари се отнасят скептично към такъв способ на въвеждане, защото може да създаде условия за нарушаване в съсирването на кръвта и развитие на некрози. Това може би е валидно при ухапвания от змии, чиито отрови силно въздействат върху съсирването на кръвта.
При ухапвания от двата подвида пепелянки, съответно южния Vipera ammodytes montandoniи номинатния Vipera ammodytes ammodytes,сме прилагали въвеждане на разреден с физиологичен разтвор серум в местата на ухапванията и това води до положителни резултати. Не се наблюдават хеморагични огнища или некрози. Това може би се дължи на факта, че отровата на двата подвида пепелянки слабо влияе на кръвосъсирването. Със сигурност подобен начин може да се използва и при ухапвания на усойница (Vipera berus)
. Преди въвеждането на серума по този начин е необходимо да се приемат антихистаминови средства: кортикостероиди и адреналин.
Подобно въвежда на серума макар и да има известна ефективност, по – скоро е от експериментален характер и няма да е от голяма полза при тежки отравяне, защото по този начин не е възможно да се въвеждат по – големи количества отрова проникнала в организма.
Въвеждане на серум при пациенти алергични към конски серум (при положителна интракожна проба)
В тези случаи където необходимо вливането на големи количества серум за кратко време методът на Безредка е неприложим.
Физиологичните показатели се контролират като при шок.
Във вената на едната ръка се поставя система с 1000 ml 5% разтвор на глюкоза с разтворен 1ml адреналин. Метода за поставяне на противозмийски серум при свръхчувствителни хора е разработен от американските медици W.Wingert и J.Wainschel през 1975 г.
Във вената на другата ръка се поставя система с 5% албумин, съдържащ 1 – 2 ампули серум. Около 3 мин преди въвеждането на серума в същата се вкарват 50 – 70 мг димедрол (или алергозан, антиалерзин).
Ако се появят симптоми на свръхчувствителност системата се спира, въвеждат се 25 – 50 ml алергозан или антиалерзин. След това вливането на албумина със серума се подновява, но с по – бавна скорост.
При пациенти с положителна кожна проба често се развиват късни алергични реакции (серумна болест) след 4 – 7 дни (понякога до 24 дни). В такива случаи се предписват антихистаминови препарати (алергозан, супрастин, фенкарол и метилпреднизолон 50 мг 4 пъти дневно).
Дози (количества) серум
Противозмийският серум е единствения специфичен антидот срещу отравяне със змийска отрова, след ухапване, но дори и този антидот не следва да се използва във всички случаи.
Серум не трябва да се ползва при „сухи” ухапвания и при минимални леки отравяния.
Абсолютни противопоказания за прилагане на противозмийски серум няма.
При средни и тежки ухапвания, по – често от пепелянки и по – рядко от усойници, серумът следва да се въвежда в максимално ранни срокове, защото това снижава тежестта на отравянето.
Според повечето клинични данни серумът може да бъде ефективен до две денонощия след ухапването. При морските змии и отровните смоци след повече от няколко дни след ухапването, при Пясъчните отровници (род Echis) до 10 дни след ухапването.
При въвеждането на серум трябва да се има предвид неговите антитоксични свойства (антитоксична активност) и симптомите на отравянето.
леко ухапване с минимално отравяне
ако за 30 – 60 мин. отока не се разпространява по пострадалия крайник, например обхваща само пръста или малка част от китката, няма затруднение в дишането, болезнени подутини, гадене или повръщане, симптоми на парастезия (свръхчувствителност) около устата или други участъци по кожата, хипотония (падане на кръвното налягане), както и изменения на лабораторните показатели – серум не се въвежда.
Отравянето се квалифицира като минимално, но наблюдението продължава 4 часа. Раната се промива и ухапания крайник може да се фиксира във функционално положение. Контрола на лабораторните показатели следва да продължи 24 часа.
средно и тежко отравяне
ако в рамките на 30 – 60 минути отока прогресира, заема цялата китка или достигне до лакътя, наблюдават се някои от симптомите на общо отравяне (болезнени подутини, затруднение в дишането, понижено кръвно налягане, гадене и повръщане и други), се провежда тест за поносимост. При отрицателна проба серума се въвежда.
1 – 2 ампули се разреждат в банка 500 ml 5% разтвор на глюкоза и чрез венозна система се въвеждат за 1 час.
Ако не се наблюдават последващи симптоми на отравянето, 1 – 2 ампули се разреждат в същото съотношения и се въвеждат за час. При по – тежко отравяне отока се разпространява широко (напр. над лакътя), появяват се гадене, повръщане, падане на кръвното налягане. С първата банка глюкоза се поставят 2 – 3 ампули и по време на вливането може да се добави още една ампула. С втората банка се вливат още 3 – 4 дози. При много тежко отравяне се въвеждат 3 – 4 ампули с първата банка, а с втората 4 – 5 ампули.
Според повечето медици работили върху лечението на ухапвания от отровни змии от пречистените противозмийски серуми, могат да бъдат въведени до 40 дози, но рядко са нужни повече от 10.
Трябва ли при ухапаните от отровни змии деца да въвеждат различни количества серум?
Не дозите са едни и същи. Определено количество серум( в зависимост от антитоксичната си активност) обезврежда определено количество отрова. Няма логика да мислим че отровната змия ще инжектира по-малко отрова при ухапване на дете в сравнение с ухапване на възрастен. .Змиите не правят точна оценка. на застрашаващите ги обекти и количеството отрова отделена при ухапване зависи от много фактори но не и от точната големина на човека, който имал нещастието да е на критично разстояние в неподходящия момент.
Ако се въведе недостатъчно количество серум значителна част отровата няма да бъде обезвредена и ако остане без да бъде инактивирана тя ще инактивира пострадалия,особено ако той е малко дете.
При всички пациенти които се лекуват с противозмийски серум, за втори път или са работили със суха змийска отрова, е препоръчително да се постави и втора система с адреналин (0,1% разтвор 1 ml), разтворен в 1000 ml 5%разтвор на глюкоза. Подкожно може да се постави ефортил. Необходимо е да има алергозан или антиалерзин (ампули) за лечение на евентуален анафилактичен шок.
снимки: Марко Иванов, Красимир Христов
Л и т е р а т у р а:
- Васильев, Д., С. Кудрявцев, О. Шумаков, 1997. Руководство по технике безопасност, методам работы с ядовитыми змеями в условиях зоопарка. Профилактика и лечение укусов, Anitvenim Index, Московски зоопарк, Москва, 2 – 27;
- Герасимов, Л. 1948. Добиване, титриране и прилагане на противозмийски серум в България, Тр. И-н Нар.Здраве, I, 9 – 37;
- Манахилов, Р., 1975, Проучвания върху отровата на българската пепелянка и стандартизация на противозмийски серум, Дисертация за кандидат на науките, 1 – 132;
- Пигулевский, С. В., 1966, Отряд Ophidia (Serpentes). Ядовитыe животные (зоотоксикология позвоночных), изд. Медицина, Ленинград, 19 – 148;
- Славчев, Р., 1959, Върху токсичната сила на змийската отрова /Vipera ammodytes/ след престояване на температура 4 – 8°С. Трудове на НИЕМ, 313 – 320;
- Султанов, М. Н, 1963. Укусы ядовитых животных, Госмедиздат, Москва, 28 – 84;
- Христов, К., Ив. Матев, Я. Илиев, 1997. Получаване на змийска отрова от пепелянка /Vipera ammodytes/ и гюрза /Vipera lebetina/, чрез електростимулация, Животновъдни науки (Приложение 1) 332 – 336;
- Христов, Кр., 2000. Влияние на комплекса от абиотични фактори върху някои имунохимични и електрофоретични свойства на отделяните при пепелянките от типичния и южния подвид отрови. Морфология, таксономия и използване на пепелянки /Vipera ammodytes (L)/ в България, Дисертация, 111 – 114;
- Christov, K., V. Beshkov, 1999. On the subspecies morphological characteristics of the sandvipers /Vipera ammodytes/ from different location in Bulgaria, Acta Zoologica Bulgarica, 51, 2/3, 43 – 50;
- Litchfield, J., F. Wilcoxon, 1949. A simplified method of evaluating dose – effect experiments, J. Pharmacol. Exper. Therap., 95, 99 – 113;
- Sutherland, S., 1979. Antivemons: Better late than never, Med. J., Aust. Vol. 2, 813 – 815;
- Wingert, W., and J. Wainschel, 1975. Diagnosis and management of envenomation by poisonous snakes, South Med. J., 68, 1015.






































 Дължина на тялото: 71см; Размах на крилете: 157см.
Дължина на тялото: 71см; Размах на крилете: 157см.














































 Тази неголяма мечка е уникална сред своите родственици, поради основното им меню насекоми – мравки, термити и др. Бърнестата мечка обитава горските и тревисти райони на Индийския полуостров. Има дълга, пухеста козина, с удължени около рамената участъци, оцветена в черно. Рядко се срещат в канелен, червеникав цвят на козината. Отпред на гърдите имат бяло или кремаво петно в U’ или Y’ форма. Характерна е източната муцуна в по-блед цвят. Формата на муцуната, голите бърни и липсата на горни резци са адаптация за хранене с насекоми. Предните лапи са извити навътре и имат дълги, леко закривени нокти, предназначени за ровене и копаене.
Тази неголяма мечка е уникална сред своите родственици, поради основното им меню насекоми – мравки, термити и др. Бърнестата мечка обитава горските и тревисти райони на Индийския полуостров. Има дълга, пухеста козина, с удължени около рамената участъци, оцветена в черно. Рядко се срещат в канелен, червеникав цвят на козината. Отпред на гърдите имат бяло или кремаво петно в U’ или Y’ форма. Характерна е източната муцуна в по-блед цвят. Формата на муцуната, голите бърни и липсата на горни резци са адаптация за хранене с насекоми. Предните лапи са извити навътре и имат дълги, леко закривени нокти, предназначени за ровене и копаене.























 Наричана на староанглийски още бруун или бруин, което също означава кафява, в Дивия Запад, мечките гризли ги наричали Олд Ефрейм /"Old Ephraim" / или Мокасиновия Джо /"Moccasin Joe"/, латинското наименование идва от латинската дума мечка - ursus и старогръцката дума мечка - arctos. Кафявите мечки са широко разпространени в голяма част от Северна Америка и Евразия в гористи и планински региони. Този вид е национално животно на много щати в Северна Америка и страни в Европа и Азия.
Наричана на староанглийски още бруун или бруин, което също означава кафява, в Дивия Запад, мечките гризли ги наричали Олд Ефрейм /"Old Ephraim" / или Мокасиновия Джо /"Moccasin Joe"/, латинското наименование идва от латинската дума мечка - ursus и старогръцката дума мечка - arctos. Кафявите мечки са широко разпространени в голяма част от Северна Америка и Евразия в гористи и планински региони. Този вид е национално животно на много щати в Северна Америка и страни в Европа и Азия.




























 Като начало промяната стартира с промяна на езика и начина на изразяване. Тази промяна започна още с появата на първите програми за чат комуникация и навлизането на емотиконите (личицата, изписани с помощта на препинателни знаци, а вече и под формата на картинки и анимации) и съкращенията. Социалните сайтове като Туитър, които налагат лимит на броя символи в една публикация, допълнително стимулират въвеждането на специфична система за комуникация чрез съкращения. Някои популярни термини като LOL (идващо от „Laughing Out Loud” – „Смеене на глас”) водят до нови думи, които имат по-широко значение от първоначалното им. Сега LOL се използва за добавяне на шеговит или забавен тон на някое съобщение, почти като вид пунктуация. Не винаги означава буквално смях. Не малко от подрастващите, особено в англоговорящите държави, комуникират основно чрез съкращения. Вече е нормално да чуеш „Omg brb asap“ (чете се буква по буква), произтичащо от начина на писане в чата и означаващо „Oh, my god I will be right back as soon as possible“, в превод „О, господи! Ще се върна възможно най-скоро“. И това е само малък пример как интернет лексиката се превръща в самостоятелен международен език. Но прекалената употреба на абревиатури или разговорни думи не винаги е подходяща. Потребителите в социалните мрежи трябва да се научат кога е подходящото време и място за различните типове писане. Дори и Туитър да прощава за съкращенията, то по-големия брой символи във Фейсбук окуражава правилното писане. Макар че съкращенията се намъкват във Фейсбук, имейлите, блоговете и дори в печатните издания, то повечето автори знаят как да пишат правилно и често го правят.
Като начало промяната стартира с промяна на езика и начина на изразяване. Тази промяна започна още с появата на първите програми за чат комуникация и навлизането на емотиконите (личицата, изписани с помощта на препинателни знаци, а вече и под формата на картинки и анимации) и съкращенията. Социалните сайтове като Туитър, които налагат лимит на броя символи в една публикация, допълнително стимулират въвеждането на специфична система за комуникация чрез съкращения. Някои популярни термини като LOL (идващо от „Laughing Out Loud” – „Смеене на глас”) водят до нови думи, които имат по-широко значение от първоначалното им. Сега LOL се използва за добавяне на шеговит или забавен тон на някое съобщение, почти като вид пунктуация. Не винаги означава буквално смях. Не малко от подрастващите, особено в англоговорящите държави, комуникират основно чрез съкращения. Вече е нормално да чуеш „Omg brb asap“ (чете се буква по буква), произтичащо от начина на писане в чата и означаващо „Oh, my god I will be right back as soon as possible“, в превод „О, господи! Ще се върна възможно най-скоро“. И това е само малък пример как интернет лексиката се превръща в самостоятелен международен език. Но прекалената употреба на абревиатури или разговорни думи не винаги е подходяща. Потребителите в социалните мрежи трябва да се научат кога е подходящото време и място за различните типове писане. Дори и Туитър да прощава за съкращенията, то по-големия брой символи във Фейсбук окуражава правилното писане. Макар че съкращенията се намъкват във Фейсбук, имейлите, блоговете и дори в печатните издания, то повечето автори знаят как да пишат правилно и често го правят. 
























